















© 2004-2021 Thomas Jäkel

Venom
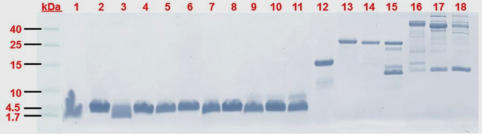
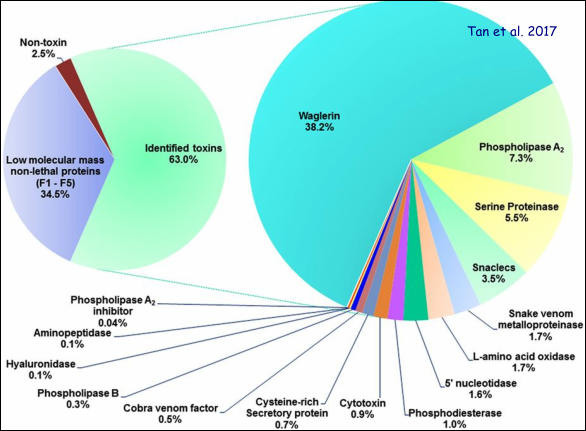
In a recent publication by Tan et al. (2017), the authors presented a comparative profiling of T. wagleri venoms (from males and females) by chromatographic and electrophoretic methods. The image above shows the result of the electrophoretic separation of two milligrams of crude T. wagleri venom after venom proteins had been fractionated by chromatographic techniques (Tan et al. 2017). Note the proteins of low molecular weight in fractions 1-11, which represent various forms of the ‘Waglerin’ toxin (neurotoxic peptides). Here, my excursion into the venomics of T. wagleri begins. Venom is a critical innovation in the evolution of advanced snakes. It represents a trophic adaptive trait, which is crucial for the foraging success of venomous snakes. Snake venoms are complex biological products, known to vary across different taxa and suited for diverse ecological niches. The pit viper T. wagleri iscategorized under Class 2 of medically important snakes by the WHO. Bites are defensive and usually uncommon, but can occur due to careless handling by keepers or disturbance brought by workers in plantations, including orchards where the snakes hunt for prey. Its bites are known to cause local envenomation (such as pain and swelling) that can be managed conservatively. Thus far, it is unclear whether this snake is able to inflict a fatal bite (Tan et al. 2017). Unlike most other pit vipers, T. wagleri venom does not possess hemorrhagic or anticoagulant activity. The venom consists of unique glycosylated phospholipase A2 and exhibits a range of common enzymatic activities: hyaluronidase, L-amino acid oxidase and phospholipase A2, but these are non-lethal in mice. The lethal components were shown to be the low molecular mass proteins of 3–8 kDa in the venom (lethal dose of approximately 0.2 μg/g in mice). These small proteins were likely the two basic and lethal polypeptides subsequently isolated from the venom, termed Waglerin I and Waglerin II. Waglerins elicit neurotoxic signs (e.g., paralysis) in laboratory animals. In the presented analysis, the waglerins accounted for almost 40% of the total venom proteins (see graph below), more than observed previously. The uniqueness of T. wagleri venom was also demonstrated by the lack of immunological recognition of its toxins by antisera or antivenoms raised against several Asiatic viperid species. The different venom properties of this species among Trimeresurus sensu lato are in line with the revised taxonomy, where it diverged earlier from the other arboreal pit vipers. Tan et al. showed that venoms from both sexes (paired peninsular and insular collections in Malaysia) demonstrated comparable lethality and protein profiles, suggesting that the venom composition of T. wagleri is relatively conserved despite its drastic sexual dimorphism. These findings imply that the venom contents of T. wagleri are consistent regardless of the sex and body size, and that predation is accomplished with the venom neurotoxic activity mediated through waglerins. Functionally, this was supported by Tan et al.’s observation of comparable lethal potency and paralytic effects of male and female venoms in mice and frogs. Mice were more susceptible than frogs to the lethal effect of both male and female venoms.



- Home
- Wagler's Viper Site (WVS)
- General Husbandry - WVS
- Breeding - WVS
- Health Problems - WVS
- Taxonomy and Phylogenetics - WVS
- Biology - WVS
- Geographic Variability - WVS
- How Females Change - WVS
- Venom - WVS
- Image Map - WVS
- Special: T. laticinctus - WVS
- Special: North Sumatra - WVS
- Special: Video on Mating - WVS
- Special: North Sulawesi - VVS
- Biological Rodent Control - BRC
- Rodent Management Laos - BRC
- Gallery
- Contact